Kombinationseignung und Heterosis
Kombinationseignung
und Heterosis![]()
1. Einleitung
1.1. Arten der Kombinationseignung
2. Feststellung der Kombinationseignung
2.1. Polycross
2.2. Topcross
2.3. M x N Kreuzungssystem
2.4. Diallele Kreuzungen
3. Diallele Kreuzungen nach Griffing
3.1. Analyse eines Griffing-Diallels,
Methode 4, Modell I
4. Beispiele
4.1. Diallel, Methode 4
4.2. M x N mating
4.3. Topcross
5. Heterosis
LITERATUR
Die Kombinationseignung bewertet das Verhalten eines Genotyps
in unterschiedlichen Kreuzungen; ein Kreuzungselter besitzt hinsichtlich
eines bestimmten Merkmals eine gute Kombinationseignung, wenn Kreuzungsnachkommenschaften
dieses Elters in ihren Merkmalsausprägungen über denen von vergleichbaren
Nachkommenschaften anderer Kreuzungseltern liegen.
Die Feststellung der Kombinationseignung erfolgt mit Hilfe von verschiedenen
Testkreuzungssystemen
und ist v.a. in der Hybridzüchtung und in der Züchtung synthetischer
Sorten zur Beurteilung von Inzuchtlinien bzw. Komponenten von besonderer
Bedeutung. Die Kombinationseignung kann auch zur Bewertung von Kreuzungskombinationen
in der Linienzüchtung bei Selbstbefruchtern verwendet werden; sie
ist hier jedoch von geringerer praktischer Bedeutung.
!!! Die Kombinationseignung eines bestimmten Genotyps ist nur durch
bestimmte Kreuzungssysteme an den Nachkommenschaften ermittelbar.
1.1. Arten der Kombinationseignung
Die Begriffe 'general combining ability' und 'specific combining ability' wurden 1942 von Sprague & Tatum eingeführt. Das Prinzip einer gezielten Linienselektion durch Nachkommenschaftsprüfungen zur züchterischen Verbesserung wurde allerdings bereits 1850 von Louis de Vilmorin (Vilmorin-Isolationsprinzip) erkannt.
(a) allgemeine Kombinationseignung (general combining ability, GCA)
Die GCA einer Linie in einem Kreuzungssystem stellt die (positive bzw. negative) Abweichung des Mittelwertes der Nachkommenschaft dieser Linie vom Gesamtmittelwert aller Nachkommenschaften des Kreuzungssystems bezüglich eines Merkmals dar.
(b) spezifische Kombinationseignung (specific combining ability, SCA)
Die SCA ist die (positive bzw. negative) Abweichung einer best. Kreuzungskombination von jenem Wert, der aufgrund der GCA-Werte der beiden Eltern für diese Kombination zu erwarten gewesen wäre.
In der Hybridzüchtung wird aus der Vielzahl möglicher Kombinationen
von zwei Inzuchtlinien jene Kombination für die Produktion von Hybridsaatgut
ausgewählt, welche sich durch die größte F1-Leistung
(Hybridleistung) und damit durch eine positive SCA der beiden Elternlinien
auszeichnet. Genetisch wird eine hohe SCA durch Dominanz-, Überdominanz-
oder andere Effekte erklärt, während die GCA einer Linie als
Ergebnis additiver Vererbung angesehen wird.
2. Feststellung der Kombinationseignung
Die Schätzung der Kombinationseignung einer Linie erfolgt anhand
bestimmter Testkreuzungen, z.B. Polycross und Topcross erlauben die Feststellung
der GCA von Genotypen; die arbeitsaufwendigeren Methoden des M x N-Kreuzungssystems
und der Diallelen Kreuzung lassen sowohl eine Schätzung der GCA als
auch der SCA zu. Um den mit Testkreuzungen verbundenen hohen Aufwand zu
reduzieren, wird neuerdings versucht, mit Hilfe molekularer Marker die
genetische
Diversität zwischen Linien zu bestimmen, um eine Vorhersage der
Heterosis bzw. Kombinationseignung zu erlangen.
... dient zur Feststellung der GCA von Zuchtmaterial (i.d.R. Klone, offen abblühende Sorten, teilweise ingezüchtete Populationen); üblich in der Populationsverbesserung und Züchtung synthetischer Sorten (Synthetics) bei fremdbestäubenden Futter- und Grünlandpflanzen.
Durchführung:
Zu prüfendes Material (nach vorangegangener Prüfung und Selektion
auf Eigenleistung) wird auf einem isolierten Polycrossfeld (Polycrossblock)
zur freien Bestäubung angebaut. Zu berücksichtigen ist, daß
nie zwei Pflanzen der gleichen Komponente nebeneinander ausgepflanzt werden
(die einzelnen Pflanzen müssen gekennzeichnet werden!; je Komponente
stehen zwischen 10 und 30 Pflanzen im Polycrossblock). Jede Komponente
soll von jeder anderen zu einem etwa gleichen Anteil bestäubt werden.
Selbstbestäubung sollte durch den Einsatz selbststeriler Formen (Selbstinkompatibilität)
unterbunden werden. Bei teilweiser Selbstfertilität sollten alle Komponenten
etwa dieselbe Rate an Selbstbefruchtung aufweisen, um die Schätzung
der GCA nicht zu verzerren.
Einzelne, gekennzeichnete Pflanzen werden geerntet, und die Samen von
allen Pflanzen der gleichen Komponenten dienen als Saatgut für eine
Leistungsprüfung in einem Feldversuch. Die aufgrund ihrer GCA besten
Klone werden in weiteren Züchtungsschritten verwendet, z.B. zur Herstellung
einer synthetischen Sorte, oder stellen zusammen eine verbesserte
Population dar.
Da mit dem Polycross-Test die GCA der Komponenten durch freie Bestäubung
jeder Komponente durch alle anderen (Pollengemisch) ermittelt wird, ist
es nötig, die einzelnen Genotypen in einer genügend großen
Anzahl an Wiederholungen anzubauen und randomisiert anzuordnen, sodaß
eine möglichst gleichmäßige Bestäubung aller Klone
durch das Pollengemisch gewährleistet ist (Voraussetzung: gleicher
Blühzeitpunkt). Für die Erhaltung der einzelnen Komponenten ist
eine vegetative Vermehrbarkeit, z.B. Stockteilung bei Gräsern, von
Vorteil.
Besteht ein Synthetic nur aus wenigen Komponenten ist die Auswahl dieser
aufgrund der GCA theoretisch nicht optimal, da es mit hoher Wahrscheinlichkeit
auch zu Paarungen zwischen Pflanzen der gleichen Komponenten kommt. Die
einzelnen Komponenten sollten auch hohe Eigenleistung besitzen, d.h. es
sollte v.a. auch eine hohe allgemeine Sorteneignung (general
varietal ability, GVA) vorhanden sein (Kombination aus GCA und Eigenleistung).
... dient zur Feststellung der GCA von Linien und wird v.a. dann eingesetzt, wenn eine große Anzahl an Genotypen zu prüfen ist, z.B. Maiszüchtung: Vielzahl an Inzucht-Linien werden mit Hilfe des Topcross auf GCA vorselektiert, bevor die Kombinationen mit der besten SCA gesucht werden.
Durchführung:
Zu prüfende I-Linien werden nicht von einem Pollengemisch, sondern
von einem speziell dazu ausgewählten Pollenspender (Tester)
bestäubt. I-Linien und Tester werden isoliert reihenweise nebeneinander
angebaut, Kastration oder Selbststerilität der I-Linien erforderlich,
damit I-Linien vom Tester bestäubt werden können (I-Linien werden
somit i.d.R. als Mutter, der/die Tester als Vater verwendet).
Das Kreuzungssaatgut wird geerntet und im darauffolgenden Jahr einer,
wenn möglich, mehrortigen Leistungsprüfung unterzogen. Die Leistung
der F1-Hybriden dient als Maß für die GCA. Die aufgrund
ihrer GCA besten Linien werden in weiteren Züchtungsschritten eingesetzt.
Bei schwer zu kastrierenden Arten kann Kreuzungssaatgut auch dadurch gewonnen
werden, daß die Pflanzen des Testers zahlenmäßig stark
in der Überzahl sind (Pollenübergewichtskreuzung).
Tester:
Entscheidend für die Effizienz des Topcross-Tests ist die Wahl
des Testers. Der Tester kann z.B. eine offen abblühende Sorte, eine
synthetische Sorte, eine I-Linie oder auch ein Hybrid sein. Wichtig ist,
daß der Tester die zu prüfenden Linien in den bedeutenden Eigenschaften
klassifizieren und differenzieren kann und nicht mit diesen verwandt ist.
Es ist zu berücksichtigen, daß ein schwacher Tester besser
zwischen Linien differenziert (etwa im Ertrag) als ein ertragreicher oder
ein an vielen Genloci dominanter, der die zu testenden Linien maskiert.
Bei Verwendung eines starken Testers wiederum, besteht die Möglichkeit,
daß bereits aus der Testkreuzung eine Hybridsorte entstehen kann.
Die Frage nach dem bestmöglichen Tester kann auf jeden Fall dahingehend
beantwortet werden, daß man nie einen einzelnen Tester verwenden
soll!
Für die erfolgreiche Durchführung des Topcross ist weiters
die gleichzeitige Blüte von Tester und zu testenden Linien erforderlich,
im Vergleich zum Polycross besteht hier jedoch die Möglichkeit, eine
gleiche Blütezeit durch gestaffelte Aussaat des Testers zu erreichen.
Was den Zeitpunkt der Durchführung des Topcross in der Maiszüchtung
betrifft, so kann man zwischen dem Testen von relativ homozygoten I-Linien
(nach vielen Inzuchtgenerationen) und dem frühen Testen (early
testing: Testung der So - oder S1 - Pflanzen)
unterscheiden.
Ein M x N-Kreuzungssystem (M x N diallel, M x N mating,
factorial) liegt dann vor, wenn eine Anzahl von m
Mutterlinien mit einer Gruppe von n Vaterlinien systematisch
gekreuzt wird, sodaß m x n Hybride entstehen. Mit Hilfe eines
solchen Kreuzungssystems ist es möglich, sowohl die GCA der Linien
als auch die SCA ganz bestimmter Kombinationen zu schätzen.
Von diallelen (od. zyklischen) Kreuzungen spricht man, wenn alle zwischen
einer bestimmten Anzahl von Linien möglichen Kreuzungskombinationen
durchgeführt und die entstandenen Hybride geprüft werden. Anhand
von solchen Diallelen können ebenfalls die GCA der einzelnen Linien
sowie die Effekte der SCA in bestimmten Kombinationen ermittelt werden.
Der Unterschied zum M x N-Design besteht darin, daß im Diallel jede
Linie sowohl Pollen- als auch Samenelter (Vater u. Mutter) ist. Von den
verbreitetsten Verfahren zur Analyse dialleler Kreuzungen soll im folgenden
auf das von Griffing näher eingegangen werden.
3. Diallele Kreuzungen nach Griffing
Grundsätzlich sind in einem Diallel von p Elternlinien
p2
Kombinationen möglich, die sich aus p Selbstungen, p*(p-1)
/ 2 Kreuzungen und p*(p-1) / 2 reziproken Kreuzungen
zusammensetzen.
Nach Griffing (1956) können vier verschiedene Methoden in Abhängigkeit
davon unterschieden werden, ob Eltern, F1 und reziproke F1
in
die Analyse einbezogen werden oder nicht:
METHODE 1 (complete diallel):
Das komplette Diallel besteht aus Eltern, F1-Hybriden und
reziproken F1-Hybriden (p2 Kombinationen)
und ermöglicht die Feststellung von reziproken Effekten und der Heterosis.
METHODE 2 (half diallel):
Eltern sowie ein Satz an F1-Hybriden werden in die Analyse
einbezogen. Es gibt somit p*(p+1) / 2 Kombinationen die eine
Feststellung der Heterosis ermöglichen.
METHODE 3:
Das Diallel setzt sich nur aus F1-Hybriden und reziproken
F1-Hybriden zusammen (p*(p-1) Kombinationen),
somit sind nur reziproke Effekte meßbar.
METHODE 4:
Nur ein Satz an F1-Hybriden (p*(p-1) / 2 Kombinationen).
Jene Methoden, welche die Eltern nicht miteinbeziehen, werden auch als
modifizierte
Diallele (modified diallels) bezeichnet.
Darüber hinaus kann zwischen zwei Modellsituationen unterschieden
werden, die sich auf die Frage beziehen, ob die Elternlinien zusammen eine
Population darstellen oder ob sie eine Zufallsstichprobe aus einer größeren
Population sind:
In der praktischen Pflanzenzüchtung, wo eine Aussage über die Kombinationseignung von bestimmten Linien angestrebt wird, geht man meist von Modell I aus. Was die anzuwendende Methode betrifft, so liefert die mit dem geringsten Kreuzungs- und Prüfungsaufwand verbundene Methode 4 bereits alle für die Hybridzüchtung erforderlichen Informationen über die Kombinationseignung der Linien. Wird jedoch das Auftreten reziproker Effekte angenommen, so kann Methode 3 angewandt werden. Da man im Rahmen der Hybridzüchtung vor allem an den F1-Hybriden und nicht an den Eltern interessiert ist, müssen letztere nicht in eine Analyse inkludiert werden, lediglich in speziellen Fällen, in denen auch die Eltern eine Rolle spielen können, z.B. bei der Züchtung synthetischer Sorten, sollten diese miteinbezogen werden.
Die praktische Durchführung einer diallelen Kreuzung geschieht in der Weise, daß zunächst alle erforderlichen Einzelkreuzungen entsprechend der gewählten Methode und mit allen in das Diallel einbezogenen Linien ausgeführt werden. Zur Auswahl der Linien sei angemerkt, daß aufgrund des großen Arbeitsaufwandes (aus 100 Linien lassen sich im geringsten Fall bereits (100 * 99) / 2 = 4950 verschiedene F1-Kombinationen herstellen) nur kleine Diallele arbeitsmäßig zu bewältigen sind, weshalb das Gros der Linien im Rahmen einer Vorselektion auf allgemeine Kombinationseignung (mittels Topcross) eliminiert werden muß.
Die aus den Kreuzungen gewonnenen F1-Hybride werden in einem
Feldversuch in mehreren Wiederholungen geprüft. Als Versuchsanlage
eignet sich eine randomisierte, vollständige Blockanlage. Grundsätzlich
sei angemerkt, daß neben F1-Hybriden auch spätere
Generationen analysiert werden können, was insbesondere dann von Vorteil
ist, wenn in der F1-Generation (v.a. bei Selbstbefruchtern)
zuwenig Saatgut für einen Feldversuch mit Wiederholungen vorhanden
ist. Der Feldversuch wird varianzanalytisch ausgewertet, die dabei festgestellte
Fehlervarianz und die Prüfgliedermittelwerte werden in einer zweiten
Varianzanalyse verwendet, um die Komponenten der Kombinationseignung schätzen
zu können.
3.1. Analyse eines Griffing-Diallels, Methode 4, Modell I
Zunächst wird die Gesamtvarianz in einen GCA-, einen SCA- und einen
Fehleranteil (wird aus der Varianzanalyse des Feldversuchs übernommen)
zerlegt und die Signifikanz dieser Varianzursachen geprüft.
Dieser Analyse liegt folgendes mathematisches Modell zugrunde:
![]()
Das bedeutet, daß der Merkmalswert xij der Kreuzung
des i-ten mit dem j-ten Elter als aus dem Gesamtmittelwert, der allgemeinen
Kombinationseignung der beiden Eltern, der spezifischen Kombinationseignung
der Kombination sowie einem Fehlereffekt zusammengesetzt gedacht wird.
Eine nicht signifikante SCA-Varianz bedeutet nach diesem Modell, daß
jede Hybridkombination aufgrund des Gesamtmittelwertes und der GCA-Werte
der Eltern bereits adäquat vorhergesagt werden kann, und die beste
Kombination entsteht, wenn die beiden Eltern mit den höchsten GCA-Werten
gekreuzt werden.
Zur Bestimmung der GCA der Linien wird für jede Linie ein Linienmittelwert
über alle F1-Hybriden, an denen diese Linie als Elter beteiligt
ist, gebildet; danach kann die GCA der Linien als positive oder negative
Abweichung des jeweiligen Linienmittelwertes vom Gesamtmittelwert errechnet
werden. Die Summe aller GCA-Werte des Diallels ergibt Null:
![]()
Um die SCA der einzelnen Kombinationen zu ermitteln, kann ein aus dem Gesamtmittelwert und den GCA-Werten der beiden Eltern zusammengesetzter Erwartungswert für den Hybrid errechnet werden; die SCA dieser Hybridkombination stellt dann die positive oder negative Abweichung des tatsächlich gemessenen Merkmalswertes vom Erwartungswert dar. Auch die Summe aller SCA-Werte über jeden Elter ergibt Null:
![]()
Die Signifikanz von Differenzen zwischen verschiedenen GCA- oder SCA-Werten
kann mit Hilfe von einfachen t-Tests ermittelt werden. Darüber
hinaus können auf Basis der Fehlervarianz aus dem Feldversuch Varianzen
von verschiedenen Werten, Effekten und Differenzen geschätzt werden.
Die Analysen von Diallelen gemäß den anderen Methoden nach
Griffing erbringen im Prinzip ähnliche Resultate; die Methoden 1 und
3 erlauben auch eine Überprüfung der Signifikanz von reziproken
Effekten.
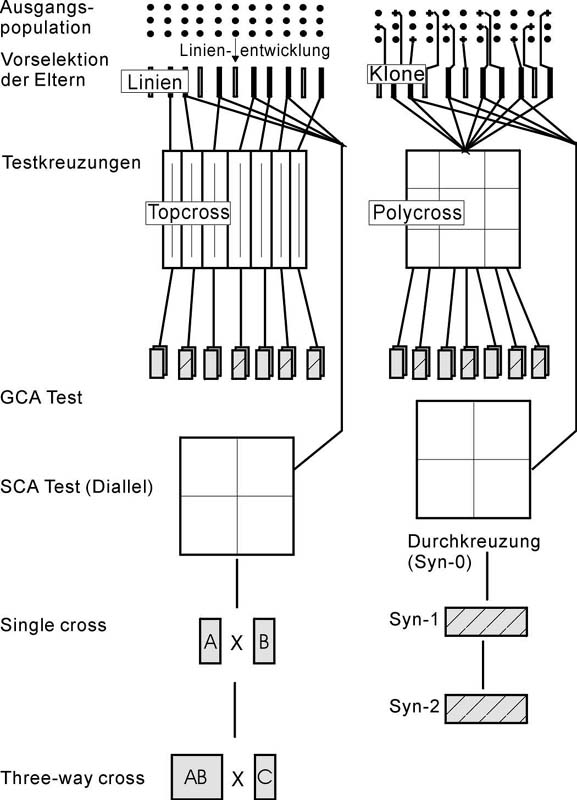
Abb. 1.: Züchtungsablauf bei der Züchtung eine Hybridsorte
(links) bzw. einer synthetischen Sorte (rechts)
| Linie B | Linie C | Linie D | S Linie | GCA Linie | GCAcorr Linie | |
| Linie A | 6.7125 | 6.39 | 5.47 | 18.5725 | 0.126 | 0.1894 |
| SCA | 0.097 | 0.147 | -0.118 | |||
| SCAcorr | -0.178 | 0.058 | 0.12 | |||
| Linie B | 6.9 | 5.855 | 19.4675 | 0.425 | 0.6369 | |
| SCA | 0.359 | -0.031 | ||||
| SCAcorr | 0.12 | 0.058 | ||||
| Linie C | 5.06 | 18.35 | 0.052 | 0.0781 | ||
| SCA | -0.454 | |||||
| SCAcorr | -0.178 | |||||
| Linie D | 16.385 | -0.603 | -0.9044 | |||
| S | 72.775 | 0.0 | 0.0 |
Sij = 36.3875
XG = 6.0646
GCAA = (SA / (n - 1)) - XG = (18.5725 / 3) - 6.0646 = 0.126
SCAAB = xAB - XG - GCAA - GCAB = 6.7125 - 6.0646 - 0.126 - 0.425 = 0.097
Korrekturfaktor: (n - 1) / (n - 2)
GCAcorrA = 0.126 * 1.5 = 0.1894
SCAcorrAB = 6.7125 - 6.0646 - 0.189 - 0.637
= -0.178
| Linie A | Linie B | Linie C | Linie D | Linie E | S Linie | GCA Linie | |
| Linie P | 10.1 | 19.7 | 20.5 | 10.7 | 10.7 | 71.7 | 0.15 |
| SCA | -0.88 | 2.25 | 1.22 | -1.48 | -1.11 | ||
| Linie Q | 14.1 | 18.4 | 22.5 | 15.1 | 12.9 | 83.0 | 2.41 |
| SCA | 0.86 | -1.31 | 0.96 | 0.66 | -1.17 | ||
| Linie R | 8.3 | 13.8 | 14.4 | 10.3 | 11.4 | 58.2 | -2.55 |
| SCA | 0.02 | -0.95 | -2.18 | 0.82 | 2.29 | ||
| S Linie | 32.5 | 51.9 | 57.4 | 36.1 | 35.0 | 212.9 | |
| GCA Linie | -3.36 | 3.11 | 4.94 | -2.16 | -2.53 |
XG = 14.1933
SCAAP = xAP - XG - GCAA
-
GCAP = 10.1 - 14.1933 - 0.15 + 3.36 = -0.88
| Inzuchtlinie | Eigenleistung | Topcross-
leistung |
GCA | Df Topcross- - Eigenleistung | Heterosis (% des besseren Elters) |
| 1 | 15 | 24 | 4.8 | 9 | 60.0 |
| 2 | 14 | 16 | -3.2 | 2 | 14.3 |
| 3 | 13 | 18 | -1.2 | 5 | 38.5 |
| 4 | 18 | 21 | 1.8 | 3 | 16.7 |
| 5 | 16 | 17 | -2.2 | 1 | 6.3 |
| Tester | 12 | ||||
| Mittelwert | 19.2 | 0.0 |
Kreuzt man zwei genetisch unterschiedliche homozygote Pflanzen miteinander, so ist die F1 i.d.R. wüchsiger und ertragreicher als das Mittel der beiden Eltern, diese Mehrleistung wird als Heterosis bezeichnet. Wird die F1 in den folgenden Generationen geselbstet, so kommt es zu einem Absinken der Leistung, der sogenannten Inzuchtdepression. Nach einer größeren Anzahl von Selbstungen sind die Pflanzen wieder weitgehend homozygot. In diesen homozygoten Linien sind die elterlichen Gene neu kombiniert, der Mittelwert aller homozygoten Linien gleicht dem Leistungsniveau der beiden ursprünglichen Eltern.
Die Höhe der Heterosis ist wesentlich vom Befruchtungssystem abhängig. Die Evolution hat bei Selbstbefruchtern immer jene Genotypen bevorzugt, die im homozygoten Zustand eine hohe Leistung erbrachten, während bei Fremdbefruchtern eine vollständige Homozygotie in natürlichen, sich zufällig paarenden Populationen nicht vorkommt. In solchen Populationen sind i.d.R. zahlreiche rezessiv vererbte Defekte verborgen, die bei Selbstung zur Inzuchtdepression führen. Im Gegenzug dazu ist die Heterosis bei Fremdbefruchtern, z.B. bei der Kreuzung zweier Inzuchtlinien, deutlich höher als bei Selbstbefruchtern. In der Folge hat sich die Hybridzüchtung v.a. bei Fremdbefruchtern (Mais, Sonnenblume, Gemüse, Roggen etc.) besonders durchgesetzt.
Die Heterosis ist i.d.R. auch umso höher, je komplexer (quantitativer) ein Merkmal vererbt wird, z.B. Ertrag. Bei Qualitätseigenschaften wie Öl- oder Proteingehalt findet man hingegen geringere Heterosis. Um eine möglichst hohe Heterosis, unabhängig vom Befruchtungssystem und der Vererbung des Merkmals, zu erreichen, sollen v.a. die beiden Eltern genetisch möglichst weit voneinander entfernt, d.h. unterschiedlich, sein.
Die Messung der Heterosis erfolgt wie oben angeführt am Elternmittel.
Man spricht von der sogenannten 'mid parent heterosis'. Züchterisch
interessant ist v.a. jedoch die Mehrleistung gegenüber dem besseren
Elter, die sogenannte 'better parent heterosis' oder Heterobeltiosis.
Niemals ist jedoch die Heterosis alleine von Interesse, sondern die Hybridleistung,
also die Summe aus Elternmittel und Heterosis.
Literatur
BAKER, R.J., 1978: Issues in diallel analysis. Crop Sci.
18:533-536
BHULLAR, G.S., GILL, K.S., KHEHRA, A.S., 1979: Combining
ability analysis over F1-F5 generations in diallel
crosses of bread wheat. Theor. Appl. Genet. 55:77-80.
CHRISTIE, B.R., SHATTUCK, V.I., 1992: The diallel cross:
design, analysis and use for plant breeders. In: Janick J (ed.), Plant
Breeding Reviews 9, 9-36. John Wiley & Sons Inc., New York.
GRIFFING, B., 1956: Concept of general and specific combining
ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 9:463-493.
HAYMAN, B.I., 1954: The analysis of variance of diallel
tables. Biometrics 10:235-244.
JINKS, J.L., 1954: The analysis of heritable variation
in a diallel cross of Nicotiana rusticana varieties. Genetics 39:767-788.
RUCKENBAUER, P., TANASCH, L., 1975: Möglichkeiten
und Grenzen dialleler Kreuzungsanalysen für die Wahl der Kreuzungseltern
in der Kreuzungszüchtung. Bericht 26. Züchtertagung, 229-241.
BAL Gumpenstein.